Accueil > Classiques > XIXe siècle > L’Origine des espèces, de Charles Darwin
Détours de la nature par la succession des générations
 L’Origine des espèces, de Charles Darwin
L’Origine des espèces, de Charles Darwin
Garnier-Flammarion, 1859, 622 p., 11,8 €.
mardi 24 novembre 2009
Le 24 novembre 1859 paraissait L’Origine des espèces, de Charles Darwin (1809-1882), un essai qui révolutionna le monde. Le 23 novembre 2009, je lis un des extraits ci-dessous à une classe de mon lycée, expliquant la teneur de ladite révolution, et la levée de boucliers opposée à Darwin. Une élève réagit en ironisant sur le thème : « il l’a bien cherché, pensez donc, ma brave dame : prétendre que les animaux n’ont pas été créés de toutes pièces par Dieu ! ». Il n’y a que deux élèves de cet acabit dans une classe de 27, mais les autres les laissent débiter leurs âneries… Actualité, donc, de Darwin ! La traduction que j’ai choisie est celle de l’édition Garnier-Flammarion mise à jour en 2008, réfection de la version originale de 1859 d’après la traduction ultérieure d’Edmond Barbier (1876), laquelle était basée sur la dernière édition du texte anglais, largement modifié par Charles Darwin. Selon Daniel Becquemont et Jean-Marc Drouin, responsables de la révision de la traduction et de la présentation de l’édition GF, ces modifications, dues surtout à la volonté de répondre à des attaques, altèrent la force de la première version. Mais comme la première traduction française de Clémence Royer (1862) est entachée de commentaires partiaux de cette dernière, Daniel Becquemont a préféré utiliser la traduction Barbier ultérieure, mais en n’en retenant que le texte originel… tout en rectifiant de nombreuses erreurs flagrantes de traduction. Cela explique donc les différences avec la version de 1906 que vous pouvez trouver sur Wikisource, à comparer avec la version anglaise de 1859 [1]. Cette lecture passionnante — et pas trop compliquée — m’a distrait de mes lectures habituelles, et rappelé feu mon bac C. Quand je pense que j’ai étudié les maths, l’anglais, la physique, et que de tout cela il ne me reste que dalle, je relativise la sacro-sainteté de mon modeste enseignement pour ces élèves qui n’en peuvent mais… Bref, j’ai surtout sélectionné quelques bonnes feuilles à exploiter dans le cadre des thèmes de culture générale en BTS 2009 et 2010 : « le Détour » et « Génération(s) ». Certains extraits sont d’ailleurs suffisamment riches pour être exploités dans les deux thèmes ! Et quand on parle de « détour », on n’oubliera pas que Darwin est passé par le détour d’un long voyage autour du monde (Patagonie, Équateur…), puis par vingt ans de réflexion et d’expérimentations, avant de publier sa théorie. On trouvera des traces de ce voyage dans Qui se souvient des hommes…, de Jean Raspail.
Un essai qui sent le soufre
En dehors du thème du BTS 2010, pour les cours au lycée, on trouvera aussi des idées, par exemple pour l’argumentation. En effet, Darwin nous propose un exemple canonique d’essai, rédigé souvent à la première personne, avec mise en œuvre d’une démarche intellectuelle qui engage l’auteur, notamment dans la voie de la négation du dogme de la création. On note aussi les longs résumés récapitulatifs à la fin de chaque chapitre, et les sous-titres indiqués en tête de chapitre. Darwin aurait-il inventé l’« abstract » ? Un fait qui me fascine est que cette thèse majeure de l’histoire des sciences et de l’humanité, contient en tout et pour tout une seule illustration, un schéma (cf. ci-dessous), auquel Darwin fait référence à maintes reprises. Belle métaphore du big bang de la réflexion humaine, qui réédite celui de la naissance du monde. Enfin, son essai est un exemple canonique de la différence entre déduction et induction. En effet, contrairement au raisonnement mathématique majoritairement déductif, Darwin utilise uniquement l’induction, c’est-à-dire qu’il base toujours ses théories sur des observations, des expériences, des faits particuliers suffisamment convaincants. Toutes ses thèses sont sujettes à vérification, et cela entraîne de fréquentes précautions oratoires.
Revenons sur la question de l’existence de Dieu. Darwin emploie souvent des périphrases et des questions rhétoriques pesées au coin du bon sens (c’est moi qui souligne) : « Pourquoi la force créatrice n’a-t-elle donc produit que des chauves-souris, à l’exclusion de tous les autres mammifères, dans les îles écartées ? » (ch. XII, p. 464). Ces pointes se multiplient dans la conclusion qui constitue le chapitre XIV (XV dans les éditions postérieures) : « s’il y a dans la nature une variabilité et un puissant agent toujours prêt à agir et à sélectionner (…) » ((ch. XIV, p. 540) ; « Au point de vue de la création indépendante de chaque être organisé et de chaque organe spécial, comment expliquer l’existence de tous ces organes portant l’empreinte la plus évidente de la plus complète inutilité (…) » (ch. XIV, p. 553) ; « Il est si facile de cacher notre ignorance sous des expressions telles que plan de création, unité de dessein, etc. (…) » ((ch. XIV, p. 553) [2]. Je citerai enfin quelques phrases-choc des deux dernières pages de la conclusion, qui constitueraient un excellent texte à étudier en classe de 1re :
« À mon avis, il me semble que ce que nous savons des lois imposées à la matière par le Créateur s’accorde mieux avec l’hypothèse que la production et l’extinction des habitants passés et présents du globe sont le résultat de causes secondaires, telles que celles qui déterminent la naissance et la mort de l’individu. Lorsque je considère tous les êtres, non plus comme des créations spéciales, mais comme les descendants en ligne directe de quelques êtres qui ont vécu longtemps avant que les premières couches du système silurien aient été déposées, ils me paraissent anoblis. À en juger d’après le passé, nous pouvons en conclure avec certitude que pas une des espèces actuellement vivantes ne transmettra sa ressemblance intacte à une époque future bien éloignée, et qu’un petit nombre d’entre elles auront seules des descendants dans les âges futurs, car le mode de groupement de tous les êtres organisés nous prouve que, dans chaque genre, le plus grand nombre des espèces, et que toutes les espèces dans beaucoup de genres, n’ont laissé aucun descendant, mais se sont totalement éteintes. Nous pouvons même jeter dans l’avenir un coup d’œil prophétique et prédire que ce sont les espèces les plus communes et les plus répandues, appartenant aux groupes les plus considérables de chaque classe, qui prévaudront ultérieurement et qui procréeront des espèces nouvelles et prépondérantes. Comme toutes les formes actuelles de la vie descendent en ligne directe de celles qui vivaient longtemps avant l’époque silurienne, nous pouvons être certains que la succession régulière des générations n’a jamais été interrompue, et qu’aucun cataclysme n’a bouleversé le monde entier. […]
Le résultat direct de cette guerre de la nature, qui se traduit par la famine et par la mort, est donc le fait le plus admirable que nous puissions concevoir, à savoir : la production des animaux supérieurs. N’y a-t-il pas une véritable grandeur dans cette manière d’envisager la vie, avec ses puissances diverses insufflées primitivement dans un petit nombre de formes [3], ou même à une seule ? Or, tandis que notre planète, obéissant à la loi fixe de la gravitation, continue à tourner dans son orbite, une quantité infinie de belles et admirables formes, sorties d’un commencement si simple, n’ont pas cessé de se développer et se développent encore ! » (p. 563).
– En dehors de ces saillies matérialistes, L’Origine des espèces n’est pas l’Almanach Vermot. C’est à peine si l’auteur se permet quelque ironie sur ses « illustres » prédécesseurs, comme le montre cet exemple le plus osé : « l’illustre Cuvier lui-même ne s’est pas aperçu qu’une balane est un crustacé, bien qu’un seul coup d’œil jeté sur la larve suffise pour ne laisser aucun doute à cet égard. » (ch XIII, p. 512). Pour conclure, rappelons que Darwin n’est en rien responsable des dévoiements de ses théories connues sous le nom de darwinisme social. Sur ce sujet, voir l’entrevue de Patrick Tort sur evene.fr, et on trouvera facilement ici ou là d’autres interventions de Patrick Tort sur la question de L’éthique darwinienne de l’altruisme.
– Pour les extraits ci-dessous, J’indique entre crochets et en gras en tête de chaque extrait en quoi il se rapporte à ces thèmes. Certains titres sont de moi ; d’autres sont inclus dans le texte ; dans ce cas, je les recopie en capitales, comme dans l’édition GF.

Photo de la statue de Darwin au Museum d’histoire naturelle de Londres.
Lutte pour l’existence
[Détours du langage ; succession des générations dans la nature]
« Je dois faire remarquer que j’emploie le terme de lutte pour l’existence dans le sens général et métaphorique, ce qui implique les relations mutuelles de dépendance des êtres organisés, et, ce qui est plus important, non seulement la vie de l’individu, mais son aptitude ou sa réussite à laisser des descendants. On peut certainement affirmer que deux animaux carnivores, en temps de famine, luttent l’un contre l’autre à qui se procurera les aliments nécessaires à son existence. Mais on dit qu’une plante, au bord du désert, lutte pour l’existence contre la sécheresse, alors qu’il serait plus exact de dire que son existence dépend de l’humidité. On pourra dire plus exactement qu’une plante, qui produit annuellement un million de graines, sur lesquelles une seule, en moyenne, parvient à se développer et à mûrir à son tour, lutte avec les plantes de la même espèce, ou d’espèces différentes, qui recouvrent déjà le sol. Le gui dépend du pommier et de quelques autres arbres ; or, c’est seulement au figuré que l’on pourra dire qu’il lutte contre ces arbres, car si des parasites en trop grand nombre s’établissent sur le même arbre, ce dernier languit et meurt ; mais on peut dire que plusieurs guis, poussant ensemble sur la même branche et produisant des graines, luttent l’un avec l’autre. Comme ce sont les oiseaux qui disséminent les graines du gui, son existence dépend d’eux, et l’on pourra dire au figuré que le gui lutte avec d’autres plantes portant des fruits, car il importe à chaque plante d’amener les oiseaux à manger les fruits qu’elle produit, pour en disséminer la graine. J’emploie donc, pour plus de commodité, le terme général lutte pour l’existence, dans ces différents sens qui se confondent les uns avec les autres.
La lutte pour l’existence résulte inévitablement de la rapidité avec laquelle tous les êtres organisés tendent à se multiplier. Tout individu qui, pendant le terme naturel de sa vie, produit plusieurs œufs ou plusieurs graines, doit être détruit à quelque période de son existence, ou pendant une saison quelconque, car, autrement le principe de l’augmentation géométrique étant donné, le nombre de ses descendants deviendrait si considérable, qu’aucun pays ne pourrait les nourrir. Aussi, comme il naît plus d’individus qu’il n’en peut vivre, il doit y avoir, dans chaque cas, lutte pour l’existence, soit avec un autre individu de la même espèce, soit avec des individus d’espèces différentes, soit avec les conditions physiques de la vie. C’est la doctrine de Malthus appliquée avec une intensité beaucoup plus considérable à tout le règne animal et à tout le règne végétal, car il n’y a là ni production artificielle d’alimentation, ni restriction apportée au mariage par la prudence. Bien que quelques espèces se multiplient aujourd’hui plus ou moins rapidement, il ne peut en être de même pour toutes, car le monde ne pourrait plus les contenir.
Il n’y a aucune exception à la règle que tout être organisé se multiplie naturellement avec tant de rapidité que, s’il n’est détruit, la terre serait bientôt couverte par la descendance d’un seul couple. L’homme même, qui se reproduit si lentement, voit son nombre doublé tous les vingt-cinq ans, et, à ce taux, en moins de mille ans, il n’y aurait littéralement plus de place sur le globe pour se tenir debout. Linné a calculé que, si une plante annuelle produit seulement deux graines — et il n’y a pas de plante qui soit si peu productive — et que l’année suivante les deux jeunes plants produisent à leur tour chacun deux graines, et ainsi de suite, on arrivera en vingt ans à un million de plants. De tous les animaux connus, l’éléphant, pense-t-on, est celui qui se reproduit le plus lentement. J’ai fait quelques calculs pour estimer quel serait probablement le taux minimum de son augmentation en nombre. On peut, sans crainte de se tromper, admettre qu’il commence à se reproduire à l’âge de trente ans, et qu’il continue jusqu’à quatre-vingt-dix ; dans l’intervalle, il produit six petits. Dans 500 ans, il y aurait 15 millions d’éléphants vivants, tous descendants du premier couple. » (p. 114).
SÉLECTION SEXUELLE.
[Détours de la séduction naturelle ; succession des générations dans la nature]
À l’état domestique, certaines particularités apparaissent souvent chez l’un des sexes et deviennent héréditaires chez ce sexe ; il en est de même à l’état de nature. Il est donc possible que la sélection naturelle modifie les deux sexes relativement à des habitudes d’existence totalement différentes comme cela arrive quelquefois, ou qu’un seul sexe se modifie relativement à l’autre sexe, ce qui arrive très souvent. Cela me conduit à dire quelques mots de ce que j’ai appelé la sélection sexuelle. Cette forme de sélection ne dépend pas de la lutte pour l’existence, mais de la lutte entre les mâles, pour s’assurer la possession des femelles. Cette lutte ne se termine pas par la mort du vaincu, mais par le défaut ou par la petite quantité de descendants. La sélection sexuelle est donc moins rigoureuse que la sélection naturelle. Ordinairement, les mâles les plus vigoureux, c’est-à-dire ceux qui sont le plus aptes à occuper leur place dans la nature, laissent un plus grand nombre de descendants. Mais, dans bien des cas, la victoire ne dépend pas tant de la vigueur générale de l’individu que de la possession d’armes spéciales qui ne se trouvent que chez le mâle. Un cerf dépourvu de bois, ou un coq dépourvu d’éperons, aurait bien peu de chances de laisser de nombreux descendants. La sélection sexuelle, en permettant toujours aux vainqueurs de se reproduire, peut donner sans doute à ceux-ci un courage indomptable, des éperons plus longs, une aile plus forte pour briser la patte du concurrent, à peu près de la même manière que le brutal éleveur de coqs de combat peut améliorer la race par le choix rigoureux de ses plus beaux adultes. Je ne saurais dire jusqu’où descend cette loi de la guerre dans l’échelle de la nature. On dit que les alligators mâles se battent, mugissent, tournent en cercle, comme le font les Indiens dans leurs danses guerrières, pour s’emparer des femelles ; on a vu des saumons mâles se battre pendant des journées entières ; les cerfs volants mâles portent quelquefois la trace des blessures que leur ont faites les larges mandibules d’autres mâles. La guerre est peut-être plus terrible encore entre les mâles des animaux polygames, car ces derniers semblent pourvus d’armes spéciales. Les animaux carnivores mâles semblent déjà bien armés, et cependant la sélection sexuelle peut encore leur donner de nouveaux moyens de défense, tels que la crinière au lion, le plastron de l’épaule au vérat et la mâchoire à crochet au saumon mâle, car le bouclier peut être aussi important que la lance au point de vue de la victoire.
Chez les oiseaux, cette lutte revêt souvent un caractère plus pacifique. Tous ceux qui ont étudié ce sujet ont constaté une ardente rivalité chez les mâles de beaucoup d’espèces pour attirer les femelles par leurs chants. Les merles de roche de la Guyane, les oiseaux de paradis, et beaucoup d’autres encore, s’assemblent en troupes ; les mâles se présentent successivement ; ils étalent avec le plus grand soin, avec le plus d’effet possible, leur magnifique plumage ; ils prennent les poses les plus extraordinaires devant les femelles, simples spectatrices, qui finissent par choisir le compagnon le plus agréable. Ceux qui ont étudié avec soin les oiseaux en captivité savent que, eux aussi, sont très susceptibles de préférences et d’antipathies individuelles : ainsi, sir R. Heron a remarqué que toutes les femelles de sa volière aimaient particulièrement un certain paon panaché. Il peut paraître enfantin d’attribuer un certain effet à des moyens apparemment si faibles : il n’est impossible d’entrer ici dans tous les détails qui seraient nécessaires ; mais, si l’homme réussit à donner en peu de temps l’élégance du port et la beauté du plumage à nos coqs Bantam, d’après le type idéal que nous concevons pour cette espèce, je ne vois pas pourquoi les oiseaux femelles ne pourraient pas obtenir un résultat semblable en choisissant, pendant des milliers de générations, les mâles qui leur paraissent les plus beaux, ou ceux dont la voix est la plus mélodieuse. On peut expliquer, en partie, par l’action de la sélection sexuelle quelques lois bien connues relatives au plumage des oiseaux mâles et femelles comparé au plumage des petits, par des variations se présentant à différents âges et transmises soit aux mâles seuls, soit aux deux sexes, à l’âge correspondant ; mais l’espace nous manque pour développer ce sujet.
Je crois donc que, toutes les fois que les mâles et les femelles d’un animal quel qu’il soit ont les mêmes habitudes générales d’existence, mais qu’ils diffèrent au point de vue de la structure, de la couleur ou de l’ornementation, ces différences sont principalement dues à la sélection sexuelle ; c’est-à-dire que certains mâles ont eu, pendant une suite non interrompue de générations, quelques légers avantages sur d’autres mâles, provenant soit de leurs armes, soit de leurs moyens de défense, soit de leur beauté ou de leurs attraits, avantages qu’ils ont transmis exclusivement à leur postérité mâle. Je ne voudrais pas cependant attribuer à cette cause toutes les différences sexuelles ; nous voyons, en effet, chez nos animaux domestiques, se produire chez les mâles des particularités qui ne semblent pas avoir été augmentées par la sélection de l’homme, comme les barbes de certains pigeons voyageurs mâles et les protubérances de certains coqs. La touffe de poils sur le jabot du dindon sauvage ne saurait lui être d’aucun avantage, il est douteux même qu’elle puisse lui servir d’ornement aux yeux de la femelle ; si même cette touffe de poils avait apparu à l’état domestique, on l’aurait considérée comme une monstruosité. » (p. 140).
La sélection naturelle sur un grand nombre de générations
[Succession des générations dans la nature]
« Après la discussion qui précède, quelque résumée qu’elle soit, nous pouvons conclure que les descendants modifiés d’une espèce quelconque réussissent d’autant mieux que leur structure est plus diversifiée et qu’ils peuvent ainsi s’emparer de places occupées par d’autres êtres. Examinons maintenant comment ces avantages résultant de la divergence des caractères tendent à agir, quand ils se combinent avec la sélection naturelle et l’extinction.
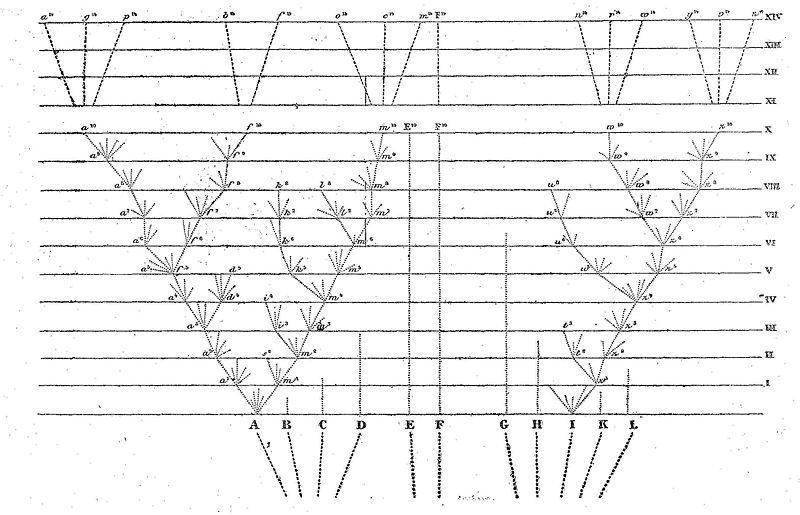
Le diagramme ci-dessus peut nous aider à comprendre ce sujet assez compliqué. Supposons que les lettres A à L représentent les espèces d’un genre riche dans le pays qu’il habite ; supposons, en outre, que ces espèces se ressemblent, à des degrés inégaux, comme cela arrive ordinairement dans la nature ; c’est ce qu’indiquent, dans le diagramme, les distances inégales qui séparent les lettres. J’ai dit un genre riche, parce que, comme nous l’avons vu dans le deuxième chapitre, plus d’espèces varient en moyenne dans un genre riche que dans un genre pauvre, et que les espèces variables des genres riches présentent un plus grand nombre de variétés. Nous avons vu aussi que les espèces les plus communes et les plus répandues varient plus que les espèces rares dont l’habitat est restreint. Supposons que A représente une espèce variable commune très répandue, appartenant à un genre riche dans son propre pays. Les lignes ponctuées divergentes, de longueur inégale, partant de A, peuvent représenter ses descendants variables. On suppose que les variations sont très légères et de la nature la plus diverse ; qu’elles ne paraissent pas toutes simultanément, mais souvent après de longs intervalles de temps, et qu’elles ne persistent pas non plus pendant des périodes égales. Les variations avantageuses seules persistent, ou, en d’autres termes, font l’objet de la sélection naturelle. C’est là que se manifeste l’importance du principe des avantages résultant de la divergence des caractères ; car ce principe détermine ordinairement les variations les plus divergentes et les plus différentes (représentées par les lignes ponctuées extérieures), que la sélection naturelle fixe et accumule. Quand une ligne ponctuée atteint une des lignes horizontales et que le point de contact est indiqué par une lettre minuscule, accompagnée d’un chiffre, on suppose qu’il s’est accumulé une quantité suffisante de variations pour former une variété bien tranchée, c’est-à-dire telle qu’on croirait devoir l’indiquer dans un ouvrage sur la zoologie systématique.
Les intervalles entre les lignes horizontales du diagramme peuvent représenter chacun mille générations ou plus. Supposons qu’après mille générations l’espèce A ait produit deux variétés bien tranchées, c’est-à-dire a1 et m1. Ces deux variétés se trouvent généralement encore placées dans des conditions analogues à celles qui ont déterminé des variations chez leurs ancêtres, d’autant que la variabilité est en elle-même héréditaire ; en conséquence, elles tendent aussi à varier, et ordinairement de la même manière que leurs ancêtres. En outre, ces deux variétés, n’étant que des formes légèrement modifiées, tendent à hériter des avantages qui ont rendu leur prototype A plus nombreux que la plupart des autres habitants du même pays ; elles participent aussi aux avantages plus généraux qui ont rendu le genre auquel appartiennent leurs ancêtres un genre riche dans son propre pays. Or, toutes ces circonstances sont favorables à la production de nouvelles variétés.
Si donc ces deux variétés sont variables, leurs variations les plus divergentes persisteront ordinairement pendant les mille générations suivantes. Après cet intervalle, on peut supposer que la variété a1 a produit la variété a2, laquelle, grâce au principe de la divergence, diffère plus de A que ne le faisait la variété a1. On peut supposer aussi que la variété m1 a produit, au bout du même laps de temps, deux variétés : m2 et s2, différant l’une de l’autre, et différant plus encore de leur souche commune A. Nous pourrions continuer à suivre ces variétés pas à pas pendant une période quelconque. Quelques variétés, après chaque série de mille générations, auront produit une seule variété, mais toujours plus modifiée ; d’autres auront produit deux ou trois variétés ; d’autres, enfin, n’en auront pas produit. Ainsi, les variétés, ou les descendants modifiés de la souche commune A, augmentent ordinairement en nombre en revêtant des caractères de plus en plus divergents. Le diagramme représente cette série jusqu’à la dix millième génération, et, sous une forme condensée et simplifiée, jusqu’à la quatorze millième.
Je ne prétends pas dire, bien entendu, que cette série soit aussi régulière qu’elle l’est dans le diagramme, bien qu’elle ait été représentée de façon assez irrégulière. Je ne prétends pas dire non plus que les variétés les plus divergentes persistent toujours ; une forme moyenne peut persister pendant longtemps et peut, ou non, produire plus d’un descendant modifié. La sélection naturelle, en effet, agit toujours en raison des places vacantes, ou de celles qui ne sont pas parfaitement occupées par d’autres êtres, et cela implique des rapports infiniment complexes. Mais, en règle générale, plus les descendants d’une espèce quelconque se modifient sous le rapport de la conformation, plus ils ont de chances de s’emparer de places et plus leur descendance modifiée tend à augmenter. Dans notre diagramme, la ligne de descendance est interrompue à des intervalles réguliers par des lettres minuscules chiffrées, indiquant les formes successives qui sont devenues suffisamment distinctes pour qu’on les reconnaisse comme variétés ; il va sans dire que ces points sont imaginaires et qu’on aurait pu les placer n’importe où, en laissant des intervalles assez longs pour permettre l’accumulation d’une somme considérable de variations divergentes.
Comme tous les descendants modifiés d’une espèce commune et très répandue, appartenant à un genre riche, tendent à participer aux avantages qui ont donné à leur ancêtre la prépondérance dans la lutte pour l’existence, ils se multiplient ordinairement en nombre, en même temps que leurs caractères deviennent plus divergents : ce fait est représenté dans le diagramme par les différentes branches divergentes partant de A. Les descendants modifiés des branches les plus récentes et les plus perfectionnées tendent à prendre la place des branches plus anciennes et moins perfectionnées, et par conséquent à les éliminer ; les branches inférieures du diagramme, qui ne parviennent pas jusqu’aux lignes horizontales supérieures, indiquent ce fait. Dans quelques cas, sans doute, les modifications portent sur une seule ligne de descendance, et le nombre des descendants modifiés ne s’accroît pas, bien que la somme des modifications divergentes ait pu augmenter. Ce cas serait représenté dans le diagramme si toutes les lignes partant de A étaient enlevées, à l’exception de celles allant de a1 à a10. Le cheval de course anglais et le limier anglais ont évidemment divergé lentement de leur souche primitive de la façon que nous venons d’indiquer, sans qu’aucun d’eux ait produit des branches ou des races nouvelles.
Supposons que, après dix mille générations, l’espèce A ait produit trois formes : a10, f10 et m10, qui, ayant divergé en caractères pendant les générations successives, en sont arrivées à différer largement, mais peut-être inégalement les unes des autres et de leur souche commune. Si nous supposons que la somme des changements entre chaque ligne horizontale du diagramme soit excessivement minime, ces trois formes ne seront encore que des variétés bien tranchées, ou bien peut-être sont-elles parvenues à la catégorie douteuse de sous-espèce ; mais nous n’avons qu’à supposer un plus grand nombre de générations, ou une modification un peu plus considérable à chaque degré, pour convertir ces trois formes en espèces bien définies. Le diagramme indique donc les degrés au moyen desquels les petites différences, séparant les variétés, s’accumulent au point de former les grandes différences séparant les espèces. En continuant la même marche un plus grand nombre de générations, ce qu’indique le diagramme sous une forme condensée et simplifiée, nous obtenons huit espèces ; a14 à m14, descendant toutes de A. C’est ainsi, je crois, que les espèces se multiplient et que les genres se forment. » (p. 171 à 177).
Les espèces distinctes présentent des variations analogues
[Tendance au retour au-delà des générations]
« Sans doute, il est très surprenant que des caractères réapparaissent après avoir disparu pendant un grand nombre de générations, des centaines peut-être. Mais, chez une race croisée une seule fois avec une autre race, la descendance présente accidentellement, pendant plusieurs générations — quelques auteurs disent pendant une douzaine ou même pendant une vingtaine —, une tendance à faire retour aux caractères de la race étrangère. Après douze générations, la proportion du sang, pour employer une expression vulgaire, de l’un des ancêtres n’est que de 1 sur 2048 ; et pourtant, comme nous le voyons, on croit généralement que cette proportion infiniment petite de sang étranger suffit à déterminer une tendance au retour. Chez une race qui n’a pas été croisée, mais chez laquelle les deux ancêtres souche ont perdu quelques caractères que possédait leur ancêtre commun, la tendance à faire retour vers ce caractère perdu pourrait, d’après tout ce que nous pouvons savoir, se transmettre de façon plus ou moins énergique pendant un nombre illimité de générations. Quand un caractère perdu reparaît chez une race après un grand nombre de générations, l’hypothèse la plus probable est, non pas que l’individu affecté se met soudain à ressembler à un ancêtre dont il est séparé par plusieurs centaines de générations, mais que le caractère en question se trouvait à l’état latent chez les individus de chaque génération successive et qu’enfin ce caractère s’est développé sous l’influence de conditions favorables, dont nous ignorons la nature. Chez les pigeons Barbe, par exemple, qui produisent très rarement des oiseaux à raies bleues et noires, il est probable qu’il y a chez les individus de chaque génération une tendance latente à la reproduction du plumage. Il ne s’agit là que d’une hypothèse, mais certains faits la corroborent, et la tendance à produire un caractère hérité pendant d’innombrables générations ne me paraît pas dans l’abstrait plus improbable que certains cas d’hérédité bien connus, d’organes tout à fait inutiles ou rudimentaires ; par exemple, chez le muflier (Antirrhinum), le rudiment d’une cinquième étamine apparaît si souvent que cette plante doit avoir une tendance héréditaire à la produire. » (p. 219).
[Je signale également le passionnant article intitulé « Instinct esclavagiste des fourmis », au sein du chapitre VII « L’instinct » (p. 281 sq. ; Attention, dans l’édition définitive, cela devient le chapitre VIII). L’édition originale comporte 14 chapitres. À partir du chapitre VIII, cela devient très scientifique et technique, et moins intéressant pour les ignares comme moi, puis à partir du chapitre XI jusqu’à la fin, c’est passionnant. Voici maintenant un extrait du chapitre XIII, où Darwin se livre à un parallèle entre langues et espèces.]
« Pour mieux faire comprendre cet exposé de la classification, prenons un exemple tiré des diverses langues humaines. Si nous possédions l’arbre généalogique complet de l’humanité, un arrangement généalogique des races humaines présenterait la meilleure classification des diverses langues parlées actuellement dans le monde entier ; si toutes les langues mortes et tous les dialectes intermédiaires et graduellement changeants devaient y être introduits, un tel groupement serait le seul possible. Cependant, il se pourrait que quelques anciennes langues, s’étant fort peu altérées, n’eussent engendré qu’un petit nombre de langues nouvelles ; tandis que d’autres, par suite de l’extension, de l’isolement, ou de l’état de civilisation des différentes races condescendantes, auraient pu se modifier considérablement et produire ainsi un grand nombre de nouveaux dialectes et de nouvelles langues. Les divers degrés de différences entre les langues dérivant d’une même souche devraient donc s’exprimer par des groupes subordonnés à d’autres groupes ; mais le seul arrangement convenable ou même possible serait encore l’ordre généalogique. Ce serait, en même temps, l’ordre strictement naturel, car il rapprocherait toutes les langues mortes et vivantes, suivant leurs affinités les plus étroites, en indiquant la filiation et l’origine de chacune d’elles. » (p. 492).
[Une autre phrase du même tonneau, au chapitre XIII]
« On peut comparer les organes rudimentaires aux lettres qui, conservées dans l’orthographe d’un mot, bien qu’inutiles pour sa prononciation, servent à en retracer l’origine et la filiation. » (p. 527).
[Un excellent résumé sur la généalogie reliant toutes les espèces.]
« En résumé, nous avons vu que la sélection naturelle, qui résulte de la lutte pour l’existence et qui implique presque inévitablement l’extinction des espèces et la divergence des caractères chez les descendants d’une même espèce parente, explique les grands traits généraux des affinités de tous les êtres organisés, c’est-à-dire leur classement en groupes subordonnés à d’autres groupes. C’est en raison des rapports généalogiques que nous classons les individus des deux sexes et de tous les âges dans une même espèce, bien qu’ils puissent n’avoir que peu de caractères en commun ; la classification des variétés reconnues, quelque différentes qu’elles soient de leurs parents, repose sur le même principe, et je crois que cet élément généalogique est le lien caché que les naturalistes ont cherché sous le nom de système naturel. Dans l’hypothèse que le système naturel, au point où il en est arrivé, est généalogique en son arrangement, les termes genres, familles, ordres, etc., n’expriment que des degrés de différence entre les descendants d’un même ancêtre et nous pouvons comprendre les règles auxquelles nous sommes forcés de nous conformer dans nos classifications. Nous pouvons comprendre pourquoi nous accordons à certaines ressemblances plus de valeur qu’à certaines autres ; pourquoi nous utilisons les organes rudimentaires et inutiles, ou n’ayant que peu d’importance physiologique ; pourquoi, en comparant un groupe avec un autre groupe distinct, nous repoussons sommairement les caractères analogues ou d’adaptation, tout en les employant dans les limites d’un même groupe. Nous voyons clairement comment il se fait que toutes les formes vivantes et éteintes peuvent être groupées en un grand système, et comment il se fait que les divers membres de chacune d’elles sont réunis les uns aux autres par les lignes d’affinité les plus complexes et les plus divergentes. Nous ne parviendrons probablement jamais à démêler l’inextricable réseau des affinités qui unissent entre eux les membres de chaque classe ; mais, si nous nous proposons un but distinct, sans chercher quelque plan de création inconnu, nous pouvons espérer faire des progrès lents, mais sûrs. » (p. 503).
– Pour terminer, quelques considérations « altersexuelles » !
Il nous faut quand même justifier la présence de cet article sur ce site. Les enseignements que tire Darwin de l’étude des hermaphrodites fourniront quelques réflexions intéressantes au point de vue altersexuel. Voici tout d’abord un extrait du chapitre IV : « La plupart des naturalistes ont dû être frappés, comme d’une étrange anomalie, du fait que, chez les animaux et chez les plantes, parmi les espèces d’une même famille et aussi d’un même genre, les unes sont hermaphrodites et les autres unisexuelles, bien qu’elles soient très semblables par tous les autres points de leur organisation. Cependant, s’il se trouve que tous les hermaphrodites se croisent de temps en temps, la différence qui existe entre eux et les espèces unisexuelles est fort insignifiante, au moins sous le rapport des fonctions. (p. 154). Le chapitre XIII fournit un contrepoint : « À l’égard des espèces à l’état de nature, chaque naturaliste a toujours fait intervenir l’élément généalogique dans ses classifications, car il comprend les deux sexes dans la dernière de ses divisions, l’espèce ; on sait, cependant, combien les deux sexes diffèrent parfois l’un de l’autre par les caractères les plus importants. C’est à peine si l’on peut attribuer un seul caractère commun aux mâles adultes et aux hermaphrodites de certains cirripèdes, que cependant personne ne songe à séparer. […] Aussitôt que l’on a reconnu que les trois formes d’orchidées (Monacanthus, Myanthus et Catasetum), antérieurement groupées sous trois genres, se rencontrent parfois sur la même plante, on les a considérées comme une seule espèce.
– Lire « Darwin, l’opiniâtre », un article de Véronique Maurus .
– Une anecdote à propos de Darwin : une citation de son récit de voyage dans le Corydon d’André Gide a fait beaucoup gloser : « J’avoue que les femmes m’ont quelque peu déçu ; elles sont loin d’être aussi belles que les hommes… ; elles gagneraient beaucoup à porter quelque vêtement. ».
– Sur les difficultés de l’enseignement de l’évolution, je signale un article de Saïda Aroua dans les Cahiers pédagogiques n° 477, décembre 2009 : « Enseigner l’évolution en Tunisie » (p. 27). Cet article ne fournit hélas pas de remède miracle, mais un regard décentré sur la Tunisie, un pays musulman où le problème se pose aussi. On regrettera une erreur manifeste : « Ce qu’a fait Darwin est une étude sous forme d’enquête et non sous forme d’expériences ». Or, au contraire, pendant les 23 années qui séparent la fin du voyage du Beagle (qui a duré 5 ans) et la publication de L’Origine des espèces, Darwin a non seulement collationné des articles et les résultats de nombreuses expériences menées par ses confrères, mais s’est livré à de nombreuses expériences personnelles, sur des espèces dont la durée de vie courte permet de tirer des conclusions sur l’évolution.
– Dans Germinal (1885), Émile Zola s’inspire de Darwin, et le mentionne.
– Erewhon, de Samuel Butler, propose une critique du darwinisme et une théorie de l’évolution alternative & dystopique.
Voir en ligne : Darwin sur Herodote.net
© altersexualite.com, 2009.
Retrouvez l’ensemble des critiques littéraires jeunesse & des critiques littéraires et cinéma adultes d’altersexualite.com. Voir aussi Déontologie critique.
[1] La meilleure version en ligne est peut-être celle de l’ABU (Association des Bibliophiles Universels), du moins permet-elle, contrairement à Wikisource, des recherches d’occurrences.
[2] Là, ce n’est pas moi qui souligne. Faut-il voir dans cette expression l’origine du gadget moderne étasunien intitulé « dessein intelligent », auquel vous me permettrez de préférer le « pastafarisme » ?!
[3] Le texte revu par l’auteur dans les éditions postérieures propose à la place de cette belle formule, l’expression : « attribuées primitivement par le Créateur à un petit nombre de formes » : bel exemple de résipiscence, et pour les cours de grammaire, d’utilisation polémique de la voix passive avec ou sans complément d’agent ! Ce seul exemple justifie l’initiative de Daniel Becquemont pour cette édition GF.